
This free science e-book is available in PDF format at:
http://www.azinet.com/aging/aging_theory_introduction.pdf
Copyright © January 2011 Azinet Press
ISBN-13 978-0-9788709-1-1
ISBN-10 0-9788709-1-3
Azinet Press
Box 239 Crownsville MD 21032
(410) 923-4745
books@azinet.com
Keywords: aging, ageing, senescence, evolution, gerontology,
geriatrics, health and fitness, bioscience, theories of aging, evolvability,
aging theories summary, regenerative medicine, adaptive aging
Cover art: Bulent Ince
Editorial assistance: Elaine Evans
Amazon Kindle edition ASIN: B004L62CEQ
Apple iBook edition: 9780978870911
Web PDF:
http://www.azinet.com/aging/aging_theory_introduction.pdf
Web epub: http://www.azinet.com/aging/aging_theory_introduction.epub
Revised: 3/21/2011
CONTENTS
Aging Mechanisms and Processes
Major Non-Programmed Aging Theories
Appendix – Evolutionary Mechanics Issues
An Introduction to Biological Aging Theory
This overview summarizes the current situation, history, major controversies, and medical implications of scientific biological aging theories. See Further Reading for a much more comprehensive treatment of this subject.
Scientific theories of biological aging (senescence) attempt to answer two questions:
How do we age? What are the specific biological mechanisms that cause aging? Aging is a very difficult subject for experimental investigation for two reasons:
First, aging is very diffuse and affects many different systems and tissues. If, for example, aging only affected the liver, we would have probably long since definitively determined the mechanisms behind aging.
Second, aging is a long-term process. An experiment to determine if a pharmaceutical agent suppresses a particular pathogen could be performed in a matter of days. An experiment to determine if an agent or protocol affects life span in mammals could take years or decades to perform.
Understanding the aging process is critical to our ability to understand and treat highly age-related diseases such as cancer and heart disease that currently kill the majority of people in developed countries, some at very young ages.
Why do we age? It is apparent that aging and life span characteristics are very specific to individual species and vary greatly between even very similar species. Mammal life spans vary over a range of about 100 to 1 between humans and the shortest-lived mouse (~0.8 years) and fish life spans vary over a range of at least 600 to 1. Some aspect of the design of each particular species therefore must determine life span. We look to evolution theory to explain why different species have different designs and evolution theory is consequently critical to attempts to explain why we age. Unfortunately, as will be described, aging and life span observations are among the very few observations that appear to conflict with Darwin’s theory and no scientific agreement has been reached regarding evolutionary explanations for aging despite 150 years of effort.
Because of the experimental difficulties, theories as to why we age are very important in providing guidance to experimental approaches. Many experimental proposals are suggested by a specific evolution-based aging theory.
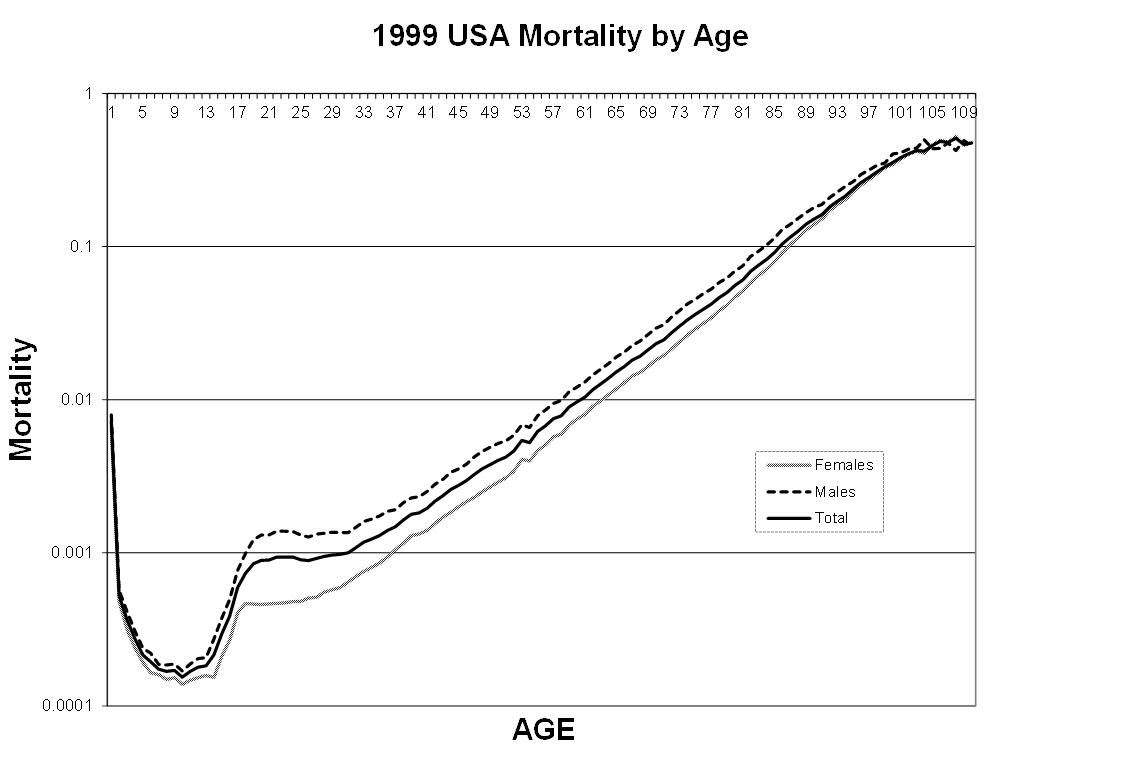
The chart below shows USA 1999 cohort death rates from all causes as a function of age at death (National Center for Health Statistics[1]). This is a log chart. The probability of death increases exponentially from about age 30, doubling approximately every seven years. In other words, aging is a major contributor to death rate starting at age 30. Aging is not just a problem for old people; about half of mortality in 37-year-olds results from aging. Curiously, death rates level off and even decline slightly for extremely old (100+) people.
Charles Darwin[2] published his book On the Origin of Species in 1859 and proposed that current organisms were descended from earlier species and further that the evolution process was directed by natural selection or “survival of the fittest.” Darwin thought that evolution was very incremental and occurred in “tiny steps.” The differences between a human and a single-cell organism were the result of accumulating these minute increments for billions of years. Note that this idea requires that the natural selection process be capable of distinguishing between very small differences in survival or reproductive capability.
It was understood that the design of an organism involves myriad compromises or tradeoffs. Strength might be a tradeoff with speed. A water buffalo might have the same ability to survive and reproduce as a gazelle.
It was widely known prior to Darwin that the design of a species could be altered by selective breeding (and by extension, natural selection). However, it was also known that no amount of selective breeding of dogs could ever create a cat. Selective breeding (and natural selection) can only alter traits that vary between interbreeding individuals. Species (essentially by definition) differ in regard to design characteristics that do not vary between individuals. Therefore, prior to Darwin, there was no apparent way that natural selection could create a new species. Darwin’s idea was that occasional inheritable mutations to individual organisms created new small variations within a species on a time scale that was so slow as to be unobservable.
There is currently no scientific disagreement regarding the idea that evolution of Earth life has occurred and the vast majority of biological observations match Darwin’s concept. Introductory biology courses currently teach that Darwin’s natural selection theory is scientifically generally accepted as the complete and comprehensive explanation for the evolution process.
However, in connection with aging and life span observations, major difficulties immediately appeared. Darwin’s idea was that random small mutational changes to organisms propagated in a population if they increased the ability of the organisms possessing them to survive and reproduce. Darwin did not suggest that the evolutionary value of survival or reproduction varied as a function of age. If an organism could survive longer and reproduce more, that was good; if it could survive and reproduce yet longer, that was even better. There was no reason to believe that the Nth descendent of a parent organism was any less important to the evolution process then the first descendent. Therefore, according to Darwin, the force of evolution was toward achieving immortality or the absence of internal limitations to life span or reproductive capability in addition to adapting to external conditions that imposed limitations on life span and reproduction.
It was immediately apparent that Darwin’s idea did not match multi-species aging and life span observations. Critics wrote Darwin[3] (c. 1859) and asked, in effect: If natural selection had been accumulatively operating for billions of years selecting longer and longer-lived individuals, why hadn’t immortality been achieved? If there was some fundamental age-dependent limitation to life span or reproduction such as a law of physics or chemistry that could not be overcome by the evolution process, why were life spans of similar species so different?
Another aspect of Darwin’s theory that is important to subsequent discussion is the idea of individual benefit. Although Darwin never used that term, subsequent interpretation of Darwin’s propagation concept seemed to logically require that individual organisms survive longer and breed more in order to propagate their individual designs. Evolved organism design characteristics including inherited behavioral traits should therefore benefit individual organisms and their direct descendents against competing members of the same species. This is the “dog eat dog” aspect of Darwinian evolutionary mechanics theory. Darwin considered that competition was fiercest between members of the same species because they, by definition, had the same requirements for food and habitat. Strict Darwinists currently believe that a design characteristic that benefits species survival or provides other more diffuse “group” benefits cannot evolve if it causes any disadvantage to the ability of individual organisms to survive and reproduce.
Since such a large proportion of observations conformed to Darwin’s idea, it was reasonable to assume that eventually we would find a conforming explanation for life span observations. This did not occur. In the intervening 150 years, theorists have proposed a number of different minor modifications to Darwin’s natural selection theory in order to accommodate aging as well as some other apparent conflicts between Darwin’s theory and observations. As described in the following sections, these modifications logically result in very different aging theories, which in turn predict dramatically different concepts for biological aging mechanisms.
Most people are under the impression that there is currently no scientific disagreement with natural selection theory (not true). Apparent discrepancies between natural selection and observations have steadily increased since Darwin. Here is a brief list of apparently conflicting observations.
Aging and life span. See above.
Altruism. Animals are observed to act in a manner not consistent with their individual best interest.
Excess age of reproductive maturity. Reproductive maturity in many animals (especially males) is delayed relative to the age plausibly required for its development, often apparently an individual disadvantage.
Mating rituals. Some mating rituals appear to represent individual disadvantage.
Biological suicide. Some instances of biological suicide present no apparent offsetting individual benefit (see octopus below).
Sexual reproduction. Sexual reproduction appears to often represent a massive individual disadvantage relative to asexual reproduction (see appendix).
Inheritance mechanisms. Many genetics discoveries raise issues with traditional natural selection theory (see appendix).
Since 1962, theorists have formally proposed a number of general adjustments to natural selection theory in response to the observed discrepancies. They all suggest that more diffuse benefits in addition to individual survival or reproductive benefit can influence the evolution process.
Group Selection. A group benefit[4] can offset an individual disadvantage.
Kin selection. Benefit to a small related group[5] can offset individual disadvantage.
Gene-oriented selection. A benefit to propagation of genes can offset individual disadvantage (e.g. R. Dawkins[6] Selfish Gene Theory)
Evolvability. A benefit to the evolution process[7] can offset individual disadvantage.
None of these alternative theories suggests that Darwinian natural selection is not the most important force behind the evolution process. They all suggest that other, more subtle and diffuse factors can also influence the evolution process. Proponents claim that these theories provide explanations for all of the listed discrepancies and defend violation of the individual benefit requirement with complex arguments often based on modern genetics discoveries (see appendix).
The primary scientific objection has historically concerned propagation. Some proponents of traditional Darwinism still contend that it is “impossible” for a characteristic to propagate and be retained in a population if it causes a net individual disadvantage, regardless of any diffuse benefit.
All of the alternative theories support the idea that a limited life span could produce a selectable evolutionary benefit and that therefore organism design characteristics that purposely limit life span could be evolved and retained. Aging theories proposing specific non-individual benefits for a design-limited life span have been proposed for most of them. Aging theories to the effect that evolved organism design features purposely limit life span in order to obtain a non-individual benefit are called programmed aging theories. The first such theory was proposed by August Weismann in 1882. Weismann thought[8] that self-limited life span or “programmed death” aided the evolution process by increasing resources available for younger and therefore minutely more evolved individuals. Since then, many other non-individual benefits for self-limited life span have been proposed6. Some theorists use the term adaptive aging to mean programmed aging. Aging is an adaptation that was selected by the evolution process because it produced an evolutionary benefit.
As will be described in the following sections, other theorists have proposed changes to Darwin’s theory that are specific to the aging/life span issue in order to support some non-programmed theories of aging in mammals.
As a further confusing factor, there is substantial religious objection to both the idea of evolution and the idea that natural selection could be a comprehensive explanation for the existence of current species, especially humans. Resulting pseudoscience proposals (creationism and intelligent design) obscure genuine scientific issues surrounding evolutionary mechanics.
The evolutionary benefit or cost of a particular organism life span is central to evolutionary theories of biological aging.
The sketch below illustrates four different scientific concepts regarding the evolutionary value of life as related to age of reproductive maturity. The benefit or cost is a measure of evolutionary force toward adapting changes in the design of an organism. Each of these concepts logically leads to an entire family of corresponding biological aging theories that in turn logically lead to particular concepts regarding the aging process.
Everybody agrees that it is beneficial for an organism to live long enough to reach reproductive maturity and that degradation due to internal limitations (aging) prior to that point would represent an evolutionary disadvantage. Further, as illustrated, life span beyond the minimum required for reproduction would be useful for organisms such as mammals and birds that need additional time to protect, nurture, or train their young. Other characteristics of specific species could affect details of the evolutionary benefit of life and therefore the shape and length of the curves below.
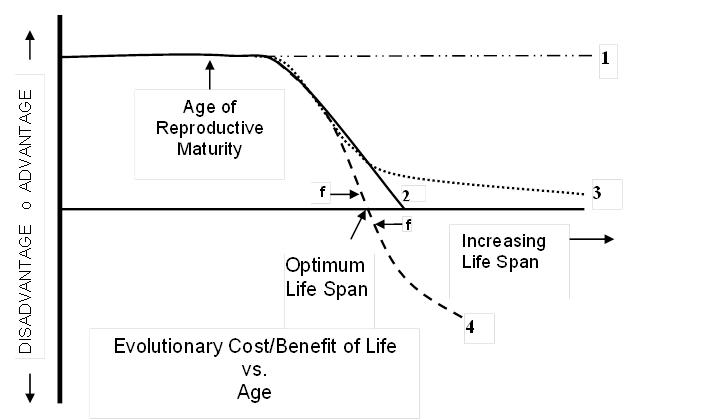
The germane scientific disagreements concern the later (older) portions of the curves during which aging occurs.
Darwin, (interrupted horizontal line 1), did not suggest that the evolutionary value of survival varied with organism age. Any incremental increase in life span added to an organism’s opportunity for reproduction and therefore created evolutionary benefit that continued indefinitely. The force of evolution was therefore toward development of immortality. Darwin’s critics immediately noted that most organisms were not immortal and that life spans varied greatly between otherwise similar organisms. Further, some plant and animal species died immediately after reproducing for the first time. These observed conflicts with Darwin’s idea eventually led to development of the other three concepts.
Peter Medawar[9] (solid line 2) proposed in 1952 that the evolutionary benefit of extended (substantially beyond age of reproductive maturity) life span in mammals becomes so negligible as to have no evolutionary effect at some species-specific age linked to reproductive maturity. Genetic drift could then introduce random changes that cause aging as long as their negative effects only occurred subsequent to that age. This led to the mutation accumulation theory of mammal aging. A yet longer life span has negligible evolutionary value but no disadvantage. His argument was that few wild mammals live long enough for aging to become a problem and the few that do have little effect on the evolution of a population. Wild organisms are subject to external limiting factors such as predators, intra-species warfare, harsh environment, lack of food supply or habitat, and infectious diseases. According to Medawar, a wild population of non-aging mammals would be very similar to a population of aging animals and would evolve in exactly the same way.
Many other proponents of non-programmed mammal aging (e.g. G. Williams[10], T. Kirkwood[11]) subsequently proposed (dotted line 3) that the evolutionary benefit of additional life span free of the deleterious effects of aging never declines to zero, essentially splitting the difference between Medawar and Darwin. A longer life would allow progressively more opportunity for reproduction and consequently at least some advantage in the propagation of an individual organism’s design. In addition, aging causes degradation at relatively young ages and this degradation has obvious negative effects on survival potential. These theorists therefore proposed that aging must be an unavoidable adverse side effect rigidly linked to some beneficial design property or properties. Because, according to this concept, the evolutionary benefit of life declines once an organism has had some opportunity to reproduce, the ultimately catastrophic disadvantage of aging could be outweighed by a relatively smaller compensating advantage to younger mammals. The assumed rigid linkage prevents the evolution process from producing a design that accomplishes the benefit without the adverse side effect. Note that both concepts 2 and 3 were developed prior to the development of any of the alternative evolutionary mechanics theories mentioned earlier.
Finally, advocates of programmed aging[12], [13], [14], [15] (dashed line 4) contend that beyond some species-specific life span, also dependent on age of reproductive maturity, additional life span creates an evolutionary disadvantage and that therefore organisms evolved mechanisms for proactively regulating their life spans to achieve an optimum life span. In this case, there would be evolutionary force (f) to both achieve the species-specific optimum life span by means of myriad evolved survival characteristics and to avoid exceeding it by means of an evolved life span regulation mechanism. Because, unlike the other concepts, there is evolutionary force toward limiting life span, there is an evolutionary rationale for the development of a complex mechanism to accomplish the limiting function. In a manner similar to many evolved mechanisms, such a regulation mechanism could include means for detecting local or temporary external conditions and optimizing an individual’s life span to fit those conditions. This concept provides a much better fit to empirical evidence than the others but requires a larger, more general, adjustment to Darwin’s idea than concepts 2 and 3.
Important Note: All of the concepts discussed here assume that reproductive decline with age is a symptom of aging not an evolutionary cause of aging. A non-aging organism would have no decline in its reproductive capability with age. If an organism had an evolved design that purposely limited its reproductive capability (e.g. otherwise unnecessary delay in reproductive maturity or purposely limited maximum reproductive age), that would present the same evolutionary issues as a design that purposely limited life span. If there was some fundamental age-dependent limitation to reproduction, why did it not apply to similar species? Some apparently non-aging organisms exist (see below) that indeed do not display either reproductive decline or decline of survival characteristics such as strength, mobility, or sensory acuity.
No one has a means for assigning any absolute value to curves 2, 3, and 4. The endless argument between proponents of these three concepts can thus be summarized: Is the evolutionary value of extended life zero, at least minutely positive, or at least minutely negative? As we will see in the next section, this hair-splitting exercise defines theories of biological aging and dramatically affects the nature of aging mechanisms predicted by the theories.
Because the differences between these value-of-life concepts (especially 2, 3, and 4) involve subtle secondary effects and complex processes operating during millions or billions of years, proving that any one of them is correct has eluded science. There is no scientific agreement regarding value-of-life. All four concepts, encompassing all of the possibilities regarding evolutionary value-of-life relative to reproductive maturity, have current followers.
There is an interesting dichotomy here: Proponents of creationism or intelligent design contend that it is “impossible” that evolutionary mechanics science will ever explain how current species came to exist and that therefore God or some other source of supernatural intelligence must have directed and implemented the design of each current species. They are not pressing similar claims about geology or astronomy, which have similar direct conflicts with religious teachings.
At the polar opposite extreme, some current theorists (see quote below) contend, in effect, that it is “impossible” that their particular current evolutionary mechanics concept (e.g. value-of-life concept 3) could be less than perfectly correct and comprehensive and that therefore any conflicting empirical evidence (or theories) must be incorrect. Claims by current scientists that multiple competing scientific theories and observations are “impossible” are essentially unheard of in other scientific disciplines. Such claims in evolutionary mechanics are likely to be at least partly a reaction to the existence of intelligent design. Recognizing that there is any scientific disagreement regarding evolutionary mechanics would result in increasing already substantial pressure to include intelligent design as a “scientific” alternative to natural selection, especially in American biology textbooks.
“The way evolution works makes it impossible for us to possess genes that are specifically designed to cause physiological decline with age or to control how long we live.” L. Hayflick, et al, Scientific American, 2004
This section summarizes four concepts regarding the processes and mechanisms that are associated with biological aging in humans and other organisms. There is relatively wide agreement that deteriorative processes that cause molecular damage are involved in the aging process. The concepts below illustrate that dramatically different aging mechanisms could exist that are all based on deteriorative processes but different value-of-life concepts. Each successive concept incorporates and is built upon the previous concept and is consequently more complex. Successive concepts provide progressively better fit to empirical evidence and additionally suggest more points at which we could attempt intervention in the aging process.
1. Simple Deterioration

Premise: Aging is simply the result of accumulative deteriorative processes such as oxidation, telomere shortening, other molecular damage, stochastic (random) changes, wear and tear, and disease-specific processes such as accumulation of cell mutations (cancer), or accumulation of blood vessel deposits or damage. Potentially many deteriorative processes are involved although some theorists believe one or another such as oxidation or telomere shortening dominates. This is the only one of the four concepts presented here that is compatible with Darwinian evolution theory and value-of-life concept 1 as taught in introductory biology classes. Consequently, most people are logically driven toward believing in simple deterioration theories. These theories tend to suggest that aging is an unalterable fact of life resulting from fundamental limitations. Billions of years of evolution that have resulted in human brains, eagle’s eyes, and other marvels of life have been unable to overcome aging.
Telomeres are “end caps” on chromosomes. Progressive shortening of telomeres during cell division has been implicated as an aging process, most notably by L. Hayflick[16] in 1961. Telomeres can be repaired by the enzyme telomerase.
Empirical Evidence: There is wide agreement that deteriorative processes exist and cause gradual deterioration in inorganic and organic systems. However, the simple deterioration concept provides a poor fit to empirical evidence. In particular, it does not explain the very large differences in life spans observed between very similar species that presumably have very similar exposure to generic deteriorative processes.
Intervention: Agents such as anti-oxidants could be sought that directly interfere with a deteriorative process. It is common practice to seek agents that interfere with disease-specific deteriorative processes such as anti-cholesterol medications.
2. Maintenance and Repair

Premise: Deteriorative processes exist but are countered and offset by maintenance and repair mechanisms whose effectiveness varies between species. The existence of these mechanisms, corresponding to the respective deteriorating processes, slows accumulation of the deteriorating effect. The effectiveness of the maintenance and repair mechanisms varies between species because evolutionary force to develop and maintain them varies according to value-of-life concept 2 or 3. Organisms with later ages of reproductive maturity needed to live longer and therefore developed and retained more effective maintenance mechanisms. Life span is not primarily limited by fundamental limitations but rather by differences in the efficiency with which different species combat deteriorative processes, an idea that increases the plausibility of intervention.
Empirical Evidence: This concept fits gradual aging and the multi-species life span variation in mammals. Additionally, we know that various maintenance mechanisms exist: hair grows, wounds heal, dead and damaged cells are replaced, and infections are combated.
Intervention: In addition to the above, we could seek agents that act to increase the effectiveness of the maintenance mechanisms, such as by increasing production of naturally occurring anti-oxidants or telomere repair enzymes.
3. Programmed Aging

Premise: Deteriorative processes exist and are offset by maintenance mechanisms but the maintenance activities are in turn modulated (attenuated) by a species-specific genetically specified biological program to result in the observed species-specific life spans. The program involves some sort of biological clock or method for determining when to slow the maintenance functions. The program and clock could be common to multiple maintenance mechanisms suggesting that signaling is involved. This idea logically descends from value-of-life concept 4. Organisms need to limit their life spans because doing so produces an evolutionary advantage according to one of the alternative evolutionary mechanics theories.
Empirical Evidence: In addition to fitting the multi-species life span observations, this concept fits discoveries of genes that cause aging in various species. It also fits observations of species such as salmon that die suddenly or age very rapidly at some point in their lives in that a program calling for that behavior is easily visualized whereas the necessarily gradual accumulation of un-repaired damage postulated in mechanism concept 2 has difficulty. Further, this concept fits observation of human genetic diseases that simultaneously cause acceleration of many (progeria) or most (Werner syndrome) symptoms of aging as these conditions could be affecting a common program controlling multiple maintenance functions.
Intervention: In addition to all of the foregoing, we could seek agents that interfere with the operation of the clock or interfere with associated signaling. Signaling in this context refers to chemical signals such as hormones that are used to coordinate biological processes within an organism or even between organisms (pheromones).
4. Regulated Programmed Aging

Premise: Deteriorative processes exist and are offset by maintenance mechanisms but the maintenance activities are modulated by a genetically specified species-specific biological program, which in turn can be adjusted by sensing of external conditions that act to alter the optimum life span.
Empirical Evidence: In addition to all of the above, this concept fits observations of explicit life span regulation in various organisms (C. Kenyon[17], et al). It also fits observations that life spans are increased by external conditions that would nominally be expected to increase deterioration such as caloric restriction or stress because sensing of these conditions could be adjusting life spans. Known biological clocks are commonly adjusted by sensing of external conditions. For example, mating seasons and circadian rhythms are synchronized to planetary cycles.
Intervention: In addition to all of the above, agents and protocols could be sought that interfere with sense functions or associated signaling.
All of these aging mechanism concepts have associated evolutionary rationales that attempt to explain why the particular mechanism should have evolved or been retained in the designs of the possessing organisms. The evolutionary arguments involve evolutionary value-of-life concepts that attempt to explain why evolution would select more effective or less effective maintenance mechanisms (concept 2) or even select mechanisms that purposely limit (concept 3) or regulate (concept 4) organism life span. Concept 4 requires substantially the same evolutionary assumptions as concept 3 but provides a better match to empirical evidence.
Since all four aging mechanism concepts involve deteriorative processes, research into direct intervention with those processes (concept 1) is the least controversial. However, ignoring the other concepts despite their superior match to empirical evidence is likely to result in missing major opportunities for successful intervention in aging processes and consequent treatments for age-related diseases and conditions.
Many gerontologists believe one of the following competing non-programmed theories of aging:
The mutation accumulation theory (P. Medawar7, 1952) proposed based on value-of-life concept 2, which Medawar originated, that aging is effectively the result of myriad universal genetic diseases. Each of these only creates problems in the age regime during which there is no evolutionary value to further life and has therefore not been selected out by the evolution process. Medawar cited Huntington’s chorea as an example of an age-related genetic disease.
The antagonistic pleiotropy theory (G. Williams8, 1957) proposes based on value-of-life concept 3, which Williams originated, that mammal aging is the result of myriad adverse side effects rigidly linked to various unknown individually beneficial characteristics. If the side effects caused problems only in later life, the associated benefits to younger mammals could provide a valid tradeoff. Pleiotropy refers to the observation that a single gene often affects different and diverse phenotypic properties and that thus there exists a linkage between those properties that would be difficult for evolution to overcome, preventing the evolution process from finding a way to accomplish the benefit without the side effect. Williams concluded that based on his theory, medical interference with the aging process was “impossible.” See appendix for more on this theory. Many other theorists contend that aging is an unavoidable side effect rigidly linked to some beneficial property.
The disposable soma theory (T. Kirkwood 9, 1977) proposes that maintenance and repair of a mammal requires substantial resources and is therefore a valid tradeoff with reproduction, (which obviously requires substantial food and energy resources) based on value-of-life concept 3. Mammals consequently did not evolve the maintenance mechanisms required for longer life.
All of these mammal aging theories have been extensively criticized by proponents of programmed aging theories. They all require one of two competing aging-specific modifications to traditional Darwinism in the form of value-of-life concepts 2 or 3 but do not require one of the more general (and more recent) alternative evolutionary mechanics theories as needed by programmed aging theories based on value-of-life concept 4.
This section presents a summary of experimental and observational evidence that provides insight into aging mechanisms.
Life Span Regulation by Sensing of External Conditions
Some investigators15, [18] report instances in which life span of simple organisms is mediated or regulated by sensing of external signals. This is typical of evolved mechanisms. In mammals, major internal biological processes are often regulated by sensing of external conditions. Examples: Circadian rhythms and annual reproductive cycles synchronize bodily processes to planetary cues.
Caloric Restriction and Life Span
Extensive experimental evidence[19] confirms that mammal life spans are typically increased, as much as doubled, when food intake is restricted and that life span continues to increase all the way to semi-starvation levels. Programmed aging theorists suggest that this behavior was selected because of evolutionary benefit. The caloric restriction effect has a group benefit in enhancing the survival potential of a group under famine conditions because a population that increased its life span while reducing its reproductive activity could survive as long with less food than another population of otherwise identical animals that did not extend their life spans and therefore had to reproduce more to maintain the same population. This idea assumes that a shorter life has an evolutionary advantage but that a tradeoff between restricting life and group survival exists. Merely surviving does not take as much energy or food as reproducing. This is a proposed example of an organism modifying an evolved genetically controlled behavior in real time to fit temporary external conditions.
Non-programmed theories have difficulty explaining the caloric restriction effect. A reduction in food would presumably reduce the resources available for maintenance and repair, increasing deterioration.
Some efforts are underway to find a “caloric restriction mimetic” that would simulate the caloric restriction effect by interfering with signaling, without requiring caloric restriction.
Stress and Life Span
Experimenters have found that several forms of stress[20] in addition to caloric restriction counter-intuitively increase life spans in various organisms. For example, exercise appears to increase life span and inactivity decreases life span. Followers of programmed aging theories suggest that this is also a selectable behavior with group benefit in a manner similar to caloric restriction. If a population of animals was under heavy predation, its members would no doubt feel more stress than another population that had few predators. If such a population increased its life span, that would tend to compensate for the higher death rate caused by predation. The adapting population would therefore have a competitive advantage over a non-adapting population.
Non-programmed theories have difficulty with the stress response. Stress would presumably increase the rate at which deterioration occurred.
Aging Genes
Several experimenters[21] have reported discovering genes that limit life span in various organisms. Deleting the genes through genetic engineering has resulted in life span increases of as much as a factor of six. Operating (expressed) genes and their associated products and processes are generally accepted to be evolved features of an organism. Programmed aging proponents say aging genes are parts of evolved mechanisms that purposely limit life span. Followers of non-programmed aging theories based on value-of-life concept 3 contend that the deleted genes must all have some individually beneficial function that compensates for their individually adverse nature. To date, no such function has been found.
Cynthia Kenyon is a leading experimentalist in this area and has found aging genes, internal hormone signaling (e.g. between digestive system and aging function), and instances where a life span regulation system is mediated by detection of external signals. Valter Longo[22] has also found experimental evidence for programmed aging.
Hutchinson-Guilford Progeria and Werner Syndrome
Hutchinson-Guilford progeria[23] and Werner syndrome[24] are single-gene human genetic diseases that dramatically accelerate multiple symptoms of aging. This suggests that there are mechanisms that are common to multiple manifestations such that a single-gene malfunction could affect multiple symptoms. This fits programmed aging theories (common life span management system) better than non-programmed theories in which multiple maintenance and repair mechanisms independently evolved.
Negligible Senescence
Organisms that do not possess deterioration with age[25] are important to aging theories and aging research because they suggest that aging is not the result of some fundamental and unalterable limitation and additionally provide clues distinguishing various theories.
Senescence refers to gradual deterioration with age (aging) and is typically very obvious under protected (zoo) conditions. More specifically, senescence refers to deterioration in survival parameters such as strength, mobility, and sensory acuity, age-related increases in disease incidence and associated death rate, and decrease in reproductive capability. Death rates for humans and most animals greatly increase with age beyond reproductive maturity. Senescence refers to internal causes of deterioration and restricted life span as opposed to external causes such as pathogens, predators, harsh conditions, and lack or food or habitat.
A very few species exhibit negligible senescence (NS). Theorists consider an organism negligibly senescent if it does not exhibit any measurable decline in survival characteristics such as strength or mobility with age, does not have a gradually increasing death rate with age, and in addition does not exhibit any measurable reduction in reproductive ability with age. The few NS species live among a wide variety of similar senescing species.
Some examples:
The Aldebra giant tortoise has a measured maximum life span (so far) of 255 years.
The Rougheye rockfish (Sebastes aleutianus) has been measured at 205 years.
Lobsters are also believed to be negligibly senescent and even apparently have increased reproductive capacity with age.
The lake sturgeon (Acipenser fulvescens) is long-lived (152 years) and may be NS.
The naked mole rat (Heterocephalus glaberis) is the only one of approximately 5500 mammal species believed to exhibit NS. These approximately mouse-size (35 grams) rodents have been observed to live 28 years vs. 1-3 years for similarly sized rodents and longer than any other rodent. Cancer has not been observed in this species.
Some clams such as Panopea generosa have long lives (~160 years) and may be NS.
The oldest known single living organism is the “Methuselah Tree”, a bristlecone pine, located in California and currently 4842 years old.
Organisms that do not age or age immeasurably slowly still die of external causes such as predator attack, accident, starvation, exposure to adverse environmental conditions, and infectious diseases. Extremely old specimens are therefore extremely rare. In some cases, measuring the age of a caught wild specimen requires killing the animal in order to measure age marks (similar to tree rings) on internal bones. We therefore have no way of knowing the maximum age that could be achieved by one of these organisms. Note that the key point with NS is lack of gradual deterioration. A hypothetical species that lived for 20 years without measurable deterioration and then died suddenly from some internal process such as semelparity would still be considered a NS species.
Although some NS species have greatly delayed sexual maturity relative to similar senescent species, others do not.
Theories to the effect that gradual deterioration is an unavoidable result of fundamental physical or chemical limitations obviously have a problem with NS. Although there are differences in metabolism between species, which could be considered differences in the rate at which the organism lives its life in a deterioration scenario, these differences are insufficient to explain the enormous differences in observed life spans, especially between species with similar metabolisms.
Semelparity and Biological Suicide
Many species of plants and animals reproduce only once (semelparity) and die suddenly after reproducing. The pink salmon dies suddenly after reproducing (at two years of age) of an apparently accelerated aging process. Some varieties of salmon survive one or two spawning periods then die suddenly following the second or third spawning showing that reproduction and associated stress is not the cause of death.
The male marsupial mice are the only semelparous mammals and die suddenly following mating at about 10 months of age.
A common explanation for semelparous behavior (compatible with value-of-life concept 3) is that suicidal behavior allows the organism to be reproductively more effective and therefore is a valid individual-benefit tradeoff with self-limited life span. In the case of the salmon, some theorists contend that the dead bodies of parents provide nutriment for their spawned immediate descendents (an individual benefit). Programmed aging proponents contend that design-limited life span has a direct evolutionary benefit.
The octopus has an especially interesting semelparous behavior. The female octopus reproduces, broods her young, and then dies of starvation. It starves because it does not eat. It does not eat because it no longer feels hunger despite its starving condition. Experiments in which the eyes were surgically removed (Wodinsky[26]) resulted in octopi that continued to eat and survive after reproducing. This demonstrates that the octopus has a complex suicide mechanism that involves connections to the nervous system to implement the behavior modification function, suggests that signaling is involved, and suggests a sense function is involved in determining when to execute the starvation behavior. This closely resembles the system described in concept 4 of the aging mechanism section. Further, the suicide of the octopus does not have any apparent individual benefit.
Evolution of Antagonistic Characteristics
Many people have a conceptual difficulty with programmed aging: How could an organism evolve myriad characteristics that obviously enable it to live longer and breed more, while simultaneously evolving characteristics that purposely limit life span (or reproduction). The key here, easily observed in nature, is that an organism can have different and conflicting requirements at different ages. For example, the North American Magicicada or 17-year locust[27] lives in the ground as a digging animal for 17 years then changes into a flying animal for a few days, reproduces, and dies. As a nymph, the cicada has zero flying ability. As an emerged adult, it has zero digging ability. The cicada is interesting in that its entire life is obviously not only “programmed” but also extremely precisely programmed. The life spans of the cicadas in a particular brood match within about 0.1 percent!
Programmed Cell Death -- Apoptosis
It is common for organisms to purposely kill their own cells (apoptosis) via a complex evolved mechanism in furtherance of growth or development tasks. Programmed organism death or phenoptosis is seen as a logical extension by proponents of programmed aging. Study of apoptosis might provide insight into aging mechanisms.
Superficial Nature of Life Span
Some characteristics of organisms vary significantly between very similar species and even between members of the same species. We think of these differences as being superficial in that they only weakly affect survival or reproductive fitness and therefore there is little natural selection force toward selecting one variation over the other. In humans, eye color apparently does not affect fitness significantly and therefore varies while eyebrows, as universal human features, are presumed to provide at least some minute survival or reproductive benefit.
Using this same logic, it is apparent that in some animals life span is superficial. Different varieties of salmon, otherwise very similar, have grossly different life spans. Other similar fish species have even more variation in life spans. Where it might appear that the shorter-lived varieties would be at a huge disadvantage that would rapidly result in their extinction, this is not the case. Apparently, if such an organism lives long enough to reach the age at which it can initially reproduce, nature does not care very much how much longer it lives.
These observations obviously conflict with the idea that life span is determined by fundamental limitations and conflict with the idea that extended life span necessarily incurs some sort of individual penalty such as reduced reproductive effectiveness or loss of some other individually beneficial function.
Some theorists favoring evolvability theories of programmed aging suggest that the disadvantage of extended life is more severe in the case of more complex organisms that display intelligence or immunity (see appendix).
Similarity of Aging Symptoms
Although there are some differences, different mammals generally share the same manifestations of aging. Cats and dogs, although having much shorter life spans share human manifestations such as cancer, heart disease, stroke, arthritis, cataracts, general weakness and loss of mobility, loss of sensory acuity, mental deficits, etc.
In connection with the maintenance and repair scenarios described earlier, one might imagine that different deteriorative processes would operate over many different time regimes. Red blood cells might die in a matter of weeks while cancer could take multiple decades to develop. Shorter-lived animals would not need to have maintenance mechanisms directed at counteracting long-term deteriorative processes.
However, if this were so we would expect to see different manifestations in different species. For example, we would not expect to see cancer in cats and dogs or shorter-lived species if it takes decades to develop. Consequently, this observation suggests that the deteriorative processes generally operate over a rather short term and that therefore all of the mammals need all of the maintenance and repair processes.
Biological Inheritance Mechanisms
All living organisms transmit digital information concerning their designs to their descendents via inheritance mechanisms such as sexual or asexual reproduction. These mechanisms are critical to the evolution process because mutational changes propagate via inheritance. Since Darwin, we have exposed enormous complexity in inheritance mechanisms much of which has plausible impact on the evolution process. Most of the previously mentioned alternative theories are either directly based on genetics discoveries (e.g. gene-centered theories) or otherwise supported by such discoveries, some rather recent. The inheritance process is still imperfectly understood. Some very recent discoveries in epigenetics[28] have the potential to radically change the way we think about biological inheritance.
Consequently, the evolution process that seemed so simple, elegant, intuitive, and certain in 1859 or even 1950 now appears to be much more complicated, messy, and uncertain.
One genetics-based concept, antagonistic pleiotropy, has been cited by proponents of both programmed and non-programmed aging theories as a justification for their positions! See appendix for more regarding effect of genetics discoveries on evolutionary mechanics theories.
Evolvability Issues
Darwin did not consider that the ability to evolve was a species-dependent variable. All living organisms were subject to mutations and natural selection. Darwin did say that natural variation in inheritable design characteristics between members of a species population was essential to the evolution process. If there were no variation, there would be nothing for natural selection to select.
Since Darwin, it has become apparent that many characteristics of particular species could affect the evolution process. Brief examples: Sexual reproduction produces more variation than asexual reproduction. Mating rituals that involve some sort of contest (e.g. Bighorn sheep) could enhance selection of characteristics that are tested by the contest. A shorter life span (beyond maturity) produces an evolvability advantage because the natural selection rate is proportional to adult death rate (see appendix). A hypothetical species that did not ever die (even from external causes) could not evolve. Other evolvability advantages of a limited life span have been proposed and some theorists12, 13 even suggest that gradual aging contributes more to evolvability than sudden death in semelparity. The ability to adapt more rapidly certainly seems to represent a competitive advantage.
The major current scientific disagreement in this area concerns whether a characteristic that produces an evolvability advantage can be selected, propagated, and retained by the evolution process if it also produces a traditional individual fitness disadvantage. This is key because evolvability characteristics generally produce fitness disadvantage or are, at best, neutral regarding individual benefit. Proponents of evolvability theories suggest evolvability explanations for all of the previously mentioned apparent discrepancies with traditional theory, and suggest solutions for the propagation issues.
Major discussions regarding evolvability are relatively recent (1995+) and post-date development of the major non-programmed aging theories. See appendix for more discussion of evolvability.
Evidence Applicability
An obvious question: Does an octopus (or worm, or fly, or fish, or even mouse) observation have any significant relevance to human aging? Many articles about human aging essentially ignore contrary evidence from non-human sources. Articles about mammal aging often ignore non-mammals while simultaneously insisting that other mammals are relevant to humans. Could a valid case be made that many of the above observations are irrelevant to human aging? The difficulty with this is that the principles underlying aging theories are extremely broad in scope. Evolution theory is specifically proposed as applying to all living organisms. The value-of-life concepts are similarly widely applicable. Mice are used as lab animals precisely because they are so biochemically similar to humans. Any suggestion that a particular observation is irrelevant should be accompanied by a plausible explanation as to why that should be the case. This is infrequently done; caviat lector.
We can define anti-aging medicine as protocols or agents that simultaneously beneficially affect two or more otherwise unrelated major manifestations of aging such as cancer and heart disease. As indicated earlier, most people are essentially trained to believe that anti-aging medicine is impossible because aging is the result of fundamental limitations. Many physicians share this view and such a view has been historically protective because of the many quacks and scammers that promote worthless aging remedies.
However, this view mediates against anti-aging research and is therefore a self-fulfilling prophecy. Attempts to find anti-aging agents are widely seen as a foolish “chase after the fountain of youth.” Few budding researchers want to embark on a career in which progress is widely seen as “impossible.”
In addition, aging is surrounded by moral, ethical, sociological, and even religious issues that do not apply to other areas of medicine. Very few people are adverse to developing treatments for or ways to prevent cancer yet informal polls indicate that as many as half of Americans have at least some issue with attempts to “treat” aging so as to extend “normal” human life span. Treating cancer is seen as extending productive life. Treating aging is seen as potentially extending the “nursing home stage” of life and “playing God.” In reality, an anti-aging agent or protocol acts to ameliorate or delay onset of age-related manifestations such as cancer. The best anti-cancer agent may well eventually turn out to be an anti-aging agent.
Public education regarding aging theory and underlying evolution theory is important because medical research is largely funded by taxes and charitable contributions.
Regardless of one’s view concerning theories of aging it is becoming increasingly clear that aging is more plastic (alterable) than widely thought. It is increasingly accepted that behavioral protocols such as exercise and caloric restriction can delay aging. There is clinical data suggesting that some agents such as aspirin and statins have a simultaneous beneficial effect on both cancer[29] and heart disease. Many items of empirical evidence previously mentioned strongly suggest aging is alterable.
A very minor improvement in human life span would have major public health impact. A ten percent increase in the life expectancy of, say, rabies patients would have little impact because so few people contract rabies and a ten percent increase in post-diagnosis life span would be insignificant. A ten percent relaxation of aging characteristics would add about six years to typical life expectancy!
Anti-Aging Research
Here are brief descriptions of a few current anti-aging research efforts:
Homo Sapiens Liberatus is an organization directed by Vladimir Skulachev, Dean of Bioengineering at Moscow State University. Recent projects include the SkQ Project to explore the use of mitochondria-targeted cationic plastoquinone derivatives (SkQs) as antioxidants specifically quenching reactive oxygen species produced by mitochondria, an event interrupting the aging program, and consequently providing treatment agents for various age-related diseases. Preliminary results in treatment of age-related eye diseases are encouraging. Skulachev is a proponent of programmed aging.
Biotime Inc. in Alameda CA (CEO Michael D. West) is investigating altering the telomere clock, telomerase therapy, and other approaches to regenerative medicine.
SENS Foundation is an organization operated by Aubrey de Grey in Cambridge, UK. Although a controversial figure, de Grey edits a journal Rejuvenation Research that attracts serious articles and has a respectable impact factor. He is a proponent of non-programmed aging.
Exeter Life Sciences is a company operated by John Sperling and Jonathan Thatcher in Phoenix AZ for research into stem cells and regenerative medicine. Their emphasis is on near-term practical applications.
Because of the ongoing scientific disagreements regarding the relationship between aging and evolution, and the simultaneous more general disagreements regarding applicable details of evolutionary mechanics, evolution-based aging theories have been notably unsuccessful in definitively explaining why we age or in aiding experimental efforts. Worse yet, despite considerable contrary evidence, many evolution-based theories either strongly imply or explicitly state that altering the aging process is “impossible” discouraging research and inhibiting funding.
· Aging remains an “unsolved problem of biology.” There is no scientific agreement regarding how or why we age.
· We cannot really understand or most effectively treat cancer or other massively age-related disease without understanding aging.
· Different theories predict dramatically different aging mechanisms and processes. They also produce very different predictions regarding the feasibility of interfering with the aging process.
· The choice of theories is largely between better fit to empirical evidence or better fit to traditional Darwinian evolutionary mechanics theory.
· Traditional Darwinism conflicts with observations other than aging resulting in multiple proposed alternatives since 1962. Our collective certainty that we completely understand the evolution process has declined. Evolutionary mechanics, an arcane area of academic science, has potentially large practical implications for public health.
· Solving the mystery of aging is within our current biotechnology capability. What is needed is the will to do so.
The book The Evolution of Aging, ISBN 0-978-87090-5 (2006) provides a much more extensive coverage of this subject.
The web site: http://www.programmed-aging.org/ provides extensive discussion of aging theories with emphasis on programmed aging and includes links to many full-text journal articles.
On the Origin of Species Charles Darwin (online free download)
http://www.literature.org/authors/darwin-charles/the-origin-of-species/
Information on negligible senescence: http://www.agelessanimals.org/
Human mortality data: http://www.mortality.org/
PubMed, operated by the U.S. National Institutes of Health, provides public online searchable catalogs including abstracts of all major journal articles concerning bioscience and has articles on all the subjects mentioned here: http://www.ncbi.nlm.nih.gov/pubmed
This appendix provides some more detail on unresolved arguments that have concerned the tiny community of evolutionary mechanics theorists for at least 50 years and summarizes a few of the many genetics discoveries that affect modern thinking about evolutionary mechanics and dependent theories of aging. As noted earlier, there are two main schools of thought: The traditional or “Darwinian” school believes that “individual” selection essentially as taught by Darwin comprehensively explains the evolution process and seeks to provide compatible explanations for apparently conflicting observations including aging. The other school believes that at least some general adjustment (such as group selection, evolvability, or gene-oriented theory) is necessary to explain observations and has produced associated explanations for the apparently discrepant observations including aging.
There is little opposition to the idea that an organism trait could produce a group or evolvability benefit. The major question is: Can an organism trait that produces a net decrease in the ability of an individual organism to survive and reproduce still propagate and be retained? Can a group or evolvability or gene-oriented benefit trade off against an individual disadvantage? If so, how? See two proposed affirmative answers below.
Evolutionary mechanics discussions tend to be rather philosophical and involve “apples and oranges” comparisons that are very difficult to quantify. Example: Would an increase in litter size and consequent improvement in reproductive capability compensate for the associated increase in maternal and offspring death rates?
Delayed Benefit Issue
All of the alternative theories (group selection, kin selection, evolvability, and gene-oriented selection) propose that a more diffuse benefit can offset an individual disadvantage. These benefits appear to require a much longer time period than individual advantage for their beneficial effect to be obtained. For example, a shorter life span would appear to be an immediate disadvantage to every organism possessing that design. Whether or not a species does or does not become extinct operates on a very much longer period. Evolvability also appears (see below) to be a long-term issue. If a species could not adapt it might still continue to exist for a long time. Is this not a very long-term issue relative to whether individuals die early? Theorists speak of the “cheater problem.” Would not an animal with a longer life span have an advantage in a population of shorter lived animals? What would keep a shorter life span from being selected out during the time required for a long-term advantage to become effective?
This suggests a crucial question: Can a long-term benefit (no matter how large) offset a short-term disadvantage (no matter how small)? Some theorists believe in “species-level group selection”, that a benefit to species survival could offset at least minor individual disadvantage; some believe only properties creating benefits to smaller groups that would become effective in a shorter time would propagate and be retained. Traditionalists reject group selection completely. Note that this takes the apples and oranges problem to a completely new and even more philosophical level: Can future apples offset present oranges? How should future benefit be discounted relative to present cost?
This question is in turn very dependent on the speed with which the evolution process is seen to operate. If the process is very slow to react then the difference between “short-term” and “long-term” would be less significant.
In this connection, most people think of selective breeding. If we took the tallest male and female dogs in the world and breed them we could produce yet taller dogs in a few generations, an eye-blink of evolutionary time. This conveys the impression that natural selection works very rapidly. The enormous anatomical differences between dogs show what can be accomplished with selective breeding in a relatively very short time.
However, there is a major difference between selective breeding and evolutionary natural selection: The breeder is attempting to enhance some specific design parameters and does not care too much about the inadvertent changes to other parameters that typically occur. Evolution “cares” about the combined net effect of all of an organism’s design parameters on survival and reproduction. This is one reason domesticated species seldom survive long in the wild.
Genetics discoveries have exposed the fact that different evolutionary processes operate over a very wide range of time periods. Let us imagine that a complex sexually-reproducing organism needs an evolutionary change. An anteater needs a longer snout because ants are building deeper nests. The evolution process needs to accomplish this change without randomly disturbing any other unrelated design parameters because any such changes are nominally adverse. The following describes the major differences in time regime that might be required.
· If the change can be accomplished by merely recombining genetic differences that already exist in the local population of a species, the change could be produced very rapidly. This is the kind of change that can be produced by selective breeding. For reasons described above, this is unlikely to satisfy the evolutionary need to produce the change without adverse changes to other parameters. In the human population, only about 0.1 percent of genetic data varies between individuals and it is unlikely that wild populations of other mammals have even that degree of variation. The other 99.9 percent cannot be altered by selective breeding or natural selection.
· If the change can be accomplished by means of a new mutation to a single gene (in addition to recombination), it would take much longer. We would need to wait until that particular mutation occurs and propagates. This is also unlikely because changing just one gene is likely to introduce adverse changes to multiple unrelated design parameters, the antagonistic pleiotropy problem.
· If the change can be accomplished by making many complementary mutational changes to many genes, it would take still longer. This is the level likely required to produce the anteater’s snout and represents the genetic differences seen between mammal species.
· If the change requires creation of a functionally new gene, we can readily imagine it might take very much longer and yet, as organisms became more complex, new genes were clearly periodically required. The creation of a functionally different new gene involves massive “chicken and egg” problems such as those associated with signals, receptors, etc. and operates on a time scale that is long relative to the time individual species exist. Genes are consequently substantially conserved between mammal species regarding their gross functionality although they vary as to details. This is the basis of the “selfish gene theory.” Gene lifetime is generally longer than species lifetime.
Linkage and Rigidity
The above discussion described a few of the different ways in which a particular organism design parameter could be linked to other design parameters. There are other ways in which linkages could occur and it is increasingly apparent that the specific design of an organism’s genome affects the path evolution follows and thus affects the evolution process.
We can use the term rigidity to describe the difficulty and therefore the time regime necessary for the evolution process to remove a linkage and allow the existence of the particular parameter without the previously linked parameter(s). How many of the above-described steps would be required for such a change? How long would it take for the evolution process to accomplish a particular change without associated linked adverse changes?
It does not appear that linkage is generally a problem within the time period a species exists. If one compares say, foot designs of various mammals, they all seem to have different designs tailored to each species’ needs. Each foot parameter seems to be independently adjustable between species. None appears to be linked to some other parameter in ways not consistent with physical requirements. Linkage did not prevent the anteater from acquiring its snout or other species from acquiring their unique characteristics.
G. Williams described in 1957 how linkage due to antagonistic pleiotropy10 between aging, a mildly individually adverse characteristic, and some unspecified individually beneficial characteristics could result in evolutionary retention of aging despite its individually adverse nature assuming value-of-life concept 3.
Precisely the same argument could be used to explain how aging, considered a mildly individually adverse but species-benefiting long-term group or evolvability characteristic could be retained despite its individually adverse nature.
However, Williams’ argument requires that the linkage be indefinitely rigid because the individual disadvantage (per concept 3) of a short life has presumably existed at least since the development of sexually reproducing organisms about 3 billion years ago. During that entire period, the evolutionary process would have been attempting to overcome the linkage and produce the benefits without the adverse side effect. Why would it not succeed?
Conversely, if aging produces a species (or shorter-term) benefit, the linkage would only need to be rigid enough to prevent aging from being selected out during a typical species lifetime, a much shorter period that is compatible with the time required to produce species-unique characteristics and the time required to overcome an antagonistic pleiotropy linkage. Therefore, an argument can be made that antagonistic pleiotropy or other causes of linkage support programmed aging via one of the alternative theories better than they support the antagonistic pleiotropy theory of non-programmed aging.
Robustness is a term commonly used to describe the ability of a particular trait to resist the effects of natural selection. Robustness regarding a particular trait could arise if that trait is controlled by multiple redundant (or partially redundant) genes. Inheritance of particular traits could be affected if they are controlled by genes on sex chromosomes (e.g. X-linkage) or in mitochondrial DNA. Because of the way the meiosis crossover mechanism operates, linkage between genes on the same chromosome is proportional to the distance (along the chromosome) between them. Transposons and other genetic mechanisms that foster movement of genetic data within a genome can affect subsequent linkages. Although the genes possessed by related species (e.g. mammals) are similar, the designs of their genomes tend to vary greatly in regard to number of chromosomes, location of genes on particular chromosomes, existence of and position of repeat patterns, introns, and other “non-functional” DNA data, and other aspects that plausibly affect linkage. There are consequently many ways in which the specific design of a particular species’ genome could affect the subsequent path of evolution in that species and thus complicate the evolution process.
Evolvability Feasibility Issues
We can define evolvability theory as the idea that a characteristic that increases an organism’s ability to evolve can be selected even if somewhat individually adverse. Evolvability is therefore a tradeoff with traditional individual benefit. As described above, many traditionalists dismiss evolvability as a form of group selection. They see evolvability as providing a very long-term benefit for the species or even future species. Some current traditionalists arguing against evolvability theories cite a 1971 book[30] written by G. Williams devoted to criticizing group selection.
However, evolvability theories (developed post-1995) appear to have logical differences from group selection that specifically affect the sort of issues described above.
When we think about the evolution process, we usually start by assuming that some phenotypic difference exists in a population such as shorter-lived and longer-lived individuals. We then analyze what would happen as time progresses. Would not the shorter-lived organisms select out prior to the time any long-term benefit would be effective? This sort of analysis is invalid for evolvability characteristics because it accounts for their disadvantages without accounting for their benefits. Evolvability characteristics benefit the evolution process by increasing local variation or otherwise increasing the effectiveness of the natural selection process. In other words, evolvability traits act to create preconditions that were necessary to create the phenotypic difference (variation) in the first place or otherwise create conditions allowing selection to occur. Without some degree of variation, the assumed phenotypic difference would not exist. Where, in group selection, we are trading a future benefit against a present disadvantage, with evolvability we are essentially trading a prior benefit against a present disadvantage. Evolvability consequently does not appear to operate in a time period longer than natural selection.
Here is a thought exercise demonstrating this issue: Imagine that a population of mice is under evolutionary pressure because of changing conditions. Between yesterday and today, this population will have minutely evolved. Proportionately more of the less fit mice will have died; slightly more of the more fit animals will have produced descendants. The size of this evolutionary increment is proportional to the amount of evolvability possessed by the population (e.g. less or more variation). In a hypothetical limit case of zero evolvability (e.g. zero variation between individuals), the increment would be zero. Therefore, evolvability affects the evolution process no matter what size time period we choose.
A case can thus be made that the mechanics of evolvability are very different from group selection and that the arguments made against group selection in 1970, even if valid, do not apply to evolvability in 2011. Any valid analysis of an evolvability trait needs to account for its benefit as well as its disadvantage.
It seems that there is little doubt that species can and do vary with regard to their capacity for further evolution. It would appear that any valid evolutionary mechanics theory would need to deal with these differences. How do these differences affect the evolution process? If they do not, why not?
Digital Genetics and “Natural” Variation
Darwin had no reason to believe that biological inheritance was not an analog process in which variation is an inherent “natural” adjunct of the process. We now know (Watson, Crick, and Franklin in 1953[31], and myriad subsequent discoveries) that inheritance is a digital communications process in which parents of any organism communicate design information to their descendents via a digital genetic code. Inheritance mechanisms must therefore conform to a series of constraints that apply to any digital communication scheme. This has implications for evolutionary mechanics as follows:
Structured, meaningful variation is not an inherent property of digital communications schemes, which tend to deliver either perfect copies or random unpredictable results. The variation we see in populations of sexually reproducing organisms is largely the result of a series of very complex and obviously evolved mechanisms that process digital inheritance data in complex ways to produce the observed structured local variation (more below). These mechanisms include paired chromosomes, meiosis, chromosome shuffling during meiosis, and unequal crossover during meiosis. The asexual reproduction mechanisms produce descendants that are much more like clones of their parent and exhibit much less variation. Local variation means variation between individuals that could plausibly be in competition with each other in a “survival of the fittest” context.
Evolution of Sexual Reproduction
Sexual reproduction, an obviously evolved and complex organism characteristic, is massively individually adverse and appears to counter basic tenets of traditional mechanics theory.
Sexually reproducing organisms developed from asexually reproducing organisms and some current organisms can reproduce either sexually or asexually. However, sexual reproduction appears to be generally individually adverse relative to asexual reproduction: Sexual reproduction produces an enormous reproductive disadvantage in that only females can produce descendants instead of all of the organisms in a population. What benefit compensated for a factor-of-two decrease in reproductive capability? Sexual reproduction entails additional risk; an individual might die before finding a mate.
If the evolutionary goal of every organism is to propagate its own personal design by living longer and breeding more, sexual reproduction appears to be a giant step backward relative to asexual reproduction. A parent organism is now sharing control over a descendant’s design with a mate and cannot assume that its descendants will reflect its personal design. Further, because of the characteristics of diploid sexual reproduction, descendants may not resemble either parent.
In a haploid organism like a bacterium, if a mutation causes a phenotypic change, every organism possessing the mutation expresses the phenotypic result. In diploid organisms that subsequently evolved, some (recessive) mutations do not result in a significant phenotypic effect unless the mutation appears in both of the organism’s genomes. This results in a situation in which mildly adverse mutations can propagate better and mildly beneficial mutations would propagate less well than in the haploid case. Why would an organism evolve a mechanism that reduces its ability to execute its evolutionary goal? One answer: More variation increases evolvability.
If variation is essential to the evolution process as specified by Darwin, sexual reproduction presents an interesting tradeoff: If an animal had an inherited behavioral trait that caused it to prefer mating with animals that most closely resembled itself, e.g. close relatives, that would produce descendants that most closely resembled the parent and thus best satisfied its evolutionary goal but exhibited little variation. If an organism had a behavioral trait that caused it to seek mates that were as different as possible from itself, that would produce more local variation at the expense of its ability to propagate its personal design. This is an example of how an evolved characteristic (the behavior) can affect the evolution process and suggests a specific tradeoff between evolvability and traditional individual benefit.
Mutations and Selectable Properties
Most people tend to think of the evolution process in terms similar to the following: A mutation occurs and causes a phenotypic change. The change either does or does not increase the organism’s ability to survive and reproduce. Natural selection either selects or does not select the change. Repeat. We can call this the “one mutation at a time” scenario.
The human genome (as determined by the Human Genome Project[32] in 2001) consists of a genetic code containing about 3.3 billion letters. Since the code uses four possible letters (A, C, G, T), each letter conveys two bits of information. The genome therefore contains 6.6 billion bits or 825 megabytes of digital data. Some of the letters are different in normal people (defined as at least one percent of the population). Perhaps the letter at position 2,323,565,022 in the code is an “A” in 12 percent of the people and a “G” in the remaining people. In the human population there are as many as ten million of these single nucleotide polymorphisms (SNPs), each of which is nominally the result of a different mutation that occurred at a different time and place. The SNPs convey most the inheritable differences between individual humans. Only a small percentage of the SNPs are in exons or otherwise likely to affect phenotype.
If we compile a list of plausible “selectable” survival parameters for a complex animal, we might include such items as speed, strength, intelligence, visual and other sensory acuity, etc. More species-specific items might include ability to climb trees, dig holes, fly long distances, etc. Each survival parameter is affected by many SNPs, each of which, individually, has little effect.
We could imagine that a complex performance parameter such as “speed” is affected by as few as 1000 of the SNPs. Each of these SNPs has a “fast” and “slow” variant. The genetically determined speed of an animal is thus determined by how many of the “fast” variants the animal possesses in its two genomes. By combining a larger number of “fast” variants, an animal might be very dramatically faster than one that possessed a large number of “slow” SNPs. From this discussion, we can see that the selectable entity is not a mutation but rather a particular combination of mutational differences that contemporaneously exist in the local population. Occasionally a new mutation introduces a new SNP to the pool.
This sort of analysis dramatically increases issues surrounding the evolution process. Instead of analyzing how a particular mutation would be processed by the inheritance mechanisms and natural selection process, we are now concerned with how combinations of mutational differences are handled during inheritance. As mentioned earlier, many aspects of inheritance mechanisms plausibly introduce linkages that affect propagation of particular combinations.
Note that sexual reproduction, by shuffling genetic data during meiosis, produces a different combination of SNPs in every individual allowing nature to rapidly test which combination produces the best evolutionary benefit. This kind of analysis strongly suggests that sexual reproduction evolved to produce local variation, an evolvability characteristic, and that an evolvability characteristic can offset an individual disadvantage.
Evolution of Intelligence
Intelligence belongs to a family of organism design characteristics that depend for their utility on the acquisition of something that accumulates during the organism’s life and consequently present a special evolvability problem. Intelligence is the ability to acquire information about the external world, store that information, and use the information to improve survival or reproductive capability. Intelligence is useless without the acquired information (experience) and conversely experience is useless without intelligence. The selectable property is therefore wisdom, essentially the product of experience and intelligence. Experience gradually accumulates during the life of an organism. If animals were immortal, the difficulty is that an older, less intelligent but more experienced animal could have more wisdom and therefore more fitness than a younger, less experienced but more intelligent animal. This situation would work against the evolution of intelligence. A design-limited life span acts to limit this otherwise destructive benefit of age. According to this concept, more complex animals that display more intelligence would obtain a larger evolvability benefit from a purposely limited life span.
Immunity presents a similar problem. The evolved characteristic here is the ability to acquire immunity to pathogen infection through progressive accumulative exposure to different pathogens. The selectable characteristic is the acquired immunity. Immortality would work against the evolution and retention of the very complex design characteristics that provide for acquisition of immunity.
Adult Death Rate and Evolvability
Adult death rate is an important evolvability parameter, especially in connection with aging and life span. To understand why this is so we need to discuss probability, performance, and expressed characteristics.
If we wanted to study how a particular coin design affected its performance in producing “heads” or “tails” in coin flipping we could perform trials. One flipping trial tells us nothing about the coin’s performance because the outcome is a matter of chance. However, if we perform very many flipping trials of coins having the same design, we could distinguish very small differences in the probability of the design producing a head vs. tail. The time required to accomplish this task is inversely proportional to the rate at which trials are performed.
In an evolution context, the life of an organism is essentially a trial of the performance of the particular design possessed by that organism regarding its ability to live longer and breed more. As with the coins, a single life has little evolutionary significance but many lives can distinguish between very small differences in the performance of a particular organism design. The rate at which organisms live their lives and therefore execute trials could be an evolvability factor in that it constrains the time required for evolution to take place. Death rate (equivalent to life rate) is therefore an evolvability factor. If organisms lived twice as long, then, everything else being equal, it would take twice as long for an evolutionary increment to take place. This idea needs further refinement as described below.
Latent characteristics, by definition, cannot affect the performance of an organism and therefore cannot affect the evolution process. In order to affect evolution the characteristic must be expressed in such a way as to plausibly affect performance of the organism. Therefore, generally speaking, deaths of juvenile organisms do not contribute to the evolution of characteristics that are only fully expressed in adults. Thus, adult death rate is an evolvability factor that affects the time required to accomplish an evolutionary increment.
Many organism design characteristics could affect adult death rate and thereby evolvability. Let us compare an immortal population to an aging mammal population. In the immortal case, some organisms would live very long lives and produce many descendents. In order for the population size to be the same as in the aging case, proportionally more animals would have to die as juveniles. There would thus be fewer adults in the immortal population. Adult death rate would be lower and the immortal population would not be able to adapt (evolve) as rapidly as the aging population. This is one of the ways in which immortality would adversely affect the evolution process.
[1] United States Centers for Disease Control (CDC) National Center for Health Statistics
[2] Charles Darwin, On the Origin of Species, 1859, ISBN 0-375-75146-7
[3] Darwin. The Origin of Species, 6th ed. 1872. Miscellaneous Objections to the Theory
[4] Wayne-Edwards V. Animal Dispersion in Relation to Social Behaviour, Edinburgh: Oliver & Boyd, 1962
[5] Hamilton, W, The Evolution of Altruistic Behavior, American Naturalist 97:354-356, 1963
[6] Dawkins R. The Selfish Gene, 1976 revised edition 1986, Oxford University Press ISBN: 0-19-286092-5
[7] Wagner G. Complex Adaptations and the Evolution of Evolvability. Evolution. 1996
[8] Weismann, August, Uber die Dauer des Lebens, Fischer, Jena
[9] Medawar P. An Unsolved Problem of Biology. 1952. H.K. Lewis & Co., London.
[10] Williams, G Pleiotropy, natural selection and the evolution of senescence. 1957. Evolution 11, 398-411
[11] Kirkwood T.B.L, Holliday F.R.S. The evolution of ageing and longevity, 1979. Proceedings of the Royal Society of London B 205: 531-546
[12] Skulachev V. Aging is a Specific Biological Function Rather than the Result of a Disorder in Complex Living Systems: Biochemical Evidence in Support of Weismann's Hypothesis. Biochemistry (Mosc). 1997 Nov;62(11):1191-5.
[13] Goldsmith T. 2008. Aging, evolvability, and the individual benefit requirement; medical implications of aging theory controversies. J. Theor. Biol. 252 (4): 764–8
[14] Mitteldorf J. Aging selected for its own sake, Evolutionary Ecology Research, 2004, 6: 1-17
[15] Libertini, G. (2008). Empirical evidence for various evolutionary hypotheses on species demonstrating increasing mortality with increasing chronological age in the wild. Scientific World Journal 19 (8): 182–93
[16] Hayflick L, Moorhead PS (1961). The serial cultivation of human diploid cell strains. Exp Cell Res 25: 585–621.
[17] Apfeld J, Kenyon C. Regulation of lifespan by sensory perception in Caenorhabditis elegans. Nature 1999.
[18] Bartke, Antebi. The endocrine regulation of aging by insulin-like signals, Science 2003 Feb 28;299(5611):1346-51
[19] Weindruch R, Walford RL, Fligiel S, Guthrie D. The retardation of aging in mice by dietary restriction: longevity, cancer, immunity and lifetime energy intake. J Nutrition 1986; 116: 641-54
[20] Liu PY, Brummel-Smith K, Ilich JZ. Aerobic exercise and whole-body vibration in offsetting bone loss in older adults. J Aging Res. 2011 Jan 3;2011:379674.
[21] Dorman J, Albinder B, Shroyer T, Kenyon C. The age-1 and daf-2 genes function in a common pathway to control the lifespan of Caenorhabditis elegans. Genetics 141(4), 1399-1406 (1995)
[22] Skulachev VP, Longo VD. Aging as a mitochondria-mediated atavistic program: can aging be switched off? Ann N Y Acad Sci. 2005 Dec;1057:145-64
[23] P Eriksson M, Brown W, Gordon L, Glynn M, Singer J, Scott L, Erdos M. Recurrent de novo point mutations in lamin A cause Hutchinson–Gilford progeria syndrome. Nature, May 2003
[24] Gray, Md; Shen, Jc; Kamath-Loeb, As; Blank, A; Sopher, Bl; Martin, Gm; Oshima, J; Loeb, La (Sep 1997). The Werner syndrome protein is a DNA helicase. Nature genetics 17 (1): 100–3. doi:10.1038/ng0997-100. PMID 9288107
[25] Buffenstein R. The naked mole-rat: a new long-living model for human aging research. J Gerontol 2005.
[26] Wodinsky J. 1977. Hormonal inhibition of feeding and death in octopus: control by optic gland secretion. Science, 198: 948–951.
[27] Williams, K. S.; Simon, C. (1995), The ecology, behavior and evolution of periodical cicadas, Annual Review of Entomology 40: 269–295
[28] O.J. Rando and K.J. Verstrepen (2007). Timescales of Genetic and Epigenetic Inheritance. Cell 128 (4): 655–668
[29] St Poynter J. et al. Statins and the Risk of Colorectal Cancer. New England Journal of Medicine; 352:2184-2192, May 26, 2005.
[30] Williams G.C. ed. 1971. Group Selection. Aldine-Atherton, Chicago
[31] Watson JD, Crick FH (April 1953). Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature 171 (4356): 737–738
[32] International Human Genome Sequencing Consortium (2001). Initial sequencing and analysis of the human genome. Nature 409 (6822): 860–921